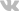
Трансдукция инсулинового сигнала

 5617
5617  0
0  Инсулин
Инсулин 24.01.2014
24.01.2014 Посттрансляционная достройка начинается в эндоплазматическом ретикулуме - гликозилирование, образование внутрицепочечных и межцепочечных -S-S-связей. Далее в аппарате Гольджи происходит протеолитическая модификация - расщепление единой пептидной цепи, образование тетрамерной молекулы, концевое гликозилирование и ацилирование жирной кислотой [Кохн К.Р., 1985]
Известно несколько субстратов рецептора инсулина (РИ): РИ-C1, РИ-С2, Shc, а также некоторые белки семейства STAT (signal transducer and activator of transcription, переносчики сигнала и активаторы транскрипции). Они активируют разные сигнальные пути. Субстрат 1 рецептора инсулина (РИ-С1) - главный. Этот цитоплазматический белок фосфорилируется по остаткам тирозина немедленно после стимуляции инсулином. Фосфорилирование субстрата РИ ведет к плейотропной реакции клетки на инсулиновый сигнал. От степени фосфорилирования субстрата зависит увеличение или уменьшение клеточного ответа на инсулин, амплитуда изменений в клетках и чувствительность к гормону. Мыши лабораторной линии, лишенные гена РИ-С1, проявляют резистентность к инсулину и сниженную толерантность при нагрузке глюкозой. Это указывает на то, что повреждения гена РИ-С1 могут быть причиной ИНЗД.
Пептидная цепь РИ-С1 содержит несколько больше 1200 аминокислотных остатков, 20 - 22 потенциальных центров фосфорилирирования по тирозину и около 40 центров Сер/Тре-фосфорилирования.
В базальном состоянии РИ-С1 фосфорилирован по серину (в меньшей мере - по треонину); после стимуляции инсулином степень фосфорилирования и по тирозину, и по серину существенно увеличивается. РИ-С1 является также и субстратом инсулиноподобного фактора роста (ИФР-I), который фосфорилирует его по тем же местам, что и РИ. Рецепторы ряда других факторов роста (например PDGF, EGF, CSF-1) не фосфорилируют РИ-С1.
Фосфорилирование РИ-С1 по нескольким тирозиновым остаткам придает ему способность соединяться с рядом белков, содержащих SH2-домены. К таким белкам в частности относятся Nck, тирозинфосфатаза syp, p85-субъединица ФИ-3-киназы, адапторный белок Grb2, протеин-тирозинфосфатаза SH-PTP2, фосфолипаза Сg, GAP (активатор малых ГТФ-связывающих белков). В результате взаимодействия РИ-С1 с подобными белками генерируются множественные нисходящие сигналы.
Нековалентное соединение белков происходит за счет взаимодействия SH2-доменов с аминокислотными последовательностями РИ-С1, содержащими фосфорилированный тирозиновый остаток. При этом не любые белки, содержащие домены SH2, присоединяются к РИ-С1: например, не присоединяются фосфолипаза Сg или GAP. Избирательность ассоциации определяется аминокислотной последовательностью в области фосфорилированного остатка тирозина (однако, домены SH2 связывают с небольшим сродством и свободный фосфотирозин). Таким путем могут образоваться многокомпонентные комплексы белков, участвующих в трансдукции сигнала. Этот механизм не является специфической особенностью инсулиновой сигнализации: при трансдукции сигналов, поступающих от рецепторов факторов роста, цитокинов и др. тоже образуются комплексы с белками, содержащими SH2-домены.
В некоторых белках, содержащих SH2-домены, а также в цитоскелетных белках найдены SH3-домены; эти домены могут взаимодействовать с пролин-богатыми последовательностями других белков.
Инсулин при посредничестве РИ-С1 активирует фосфатидилинозитол-3-киназу (ФИ-3-киназу). ФИ-3-киназа катализирует фосфорилирование ФИ, ФИ-4-Р и ФИ-4,5-Р2 по позиции 3; образуются соответственно ФИ-3-Р. ФИ-3,4-Р2 и ФИ-3,4,5-Р3.
Фермент представляет собой гетеродимер, содержащий регуляторную (р85) и каталитическую (р110) субъединицы. В регуляторной субъединице есть два SH2-домена и SH3-домен, поэтому ФИ-3-киназа с высоким сродством присоединяется к РИ-С1. Изолированные домены SH2, полученные из р85 (регуляторной субъединицы), ингибируют образование комплекса с цельной ФИ-3-киназой. Из этого следует, что фермент присоединяет РИ-С1 своей регуляторной субъединицей. При образовании комплекса ФИ-3-киназа активируется.
Активация ФИ-3-киназы является звеном сигнального пути, стимулирующего транслокацию ГЛЮТ-4 из цитозоля в плазматическую мембрану, а следовательно - и трансмембранный перенос глюкозы в мышечные и жировые клетки. Ингибиторы ФИ-3-киназы подавляют и базальное, и стимулированное инсулином потребление глюкозы; в последнем случае ингибируется транслокация ГЛЮТ-4 к мембране. В исследованиях с культурами мышечных клеток получены результаты, позволяющие предполагать следующую цепь событий при стимуляции инсулином потребления глюкозы: РИ-С1 ® ФИ-3-киназа ® ПК-С ® транслокация ГЛЮТ-4. Протеинкиназа С in vitro прямо активируется полифосфоинозитидами. Механизм активации in vivo неизвестен.
В жировых клетках активация ФИ-3-киназы инсулином приводит к ингибированию липолиза. Лимитирующей стадией липолиза в адипоцитах является реакция, катализируемая гормончувствительной липазой, которая активна в фосфорилированной форме (цАМФ-зависимое фосфорилирование). При стимуляции инсулином концентрация цАМФ в адипоцитах снижается в результате следующего каскада реакций: фосфорилируется (и активируется) протеинкиназа В (ПК-В), которая фосфорилирует (и тоже активирует) фосфодиэстеразу цАМФ:
Многие факторы роста стимулируют ФИ-3-киназу, но не влияют на обмен глюкозы. Это указывает на то, что разные сигнальные входы имеют специфические механизмы для использования ФИ-3-киназной системы, чтобы генерировать специфический конечный ответ.
Инсулин и факторы роста, действующие через Тир-киназные рецепторы, активируют ФИ-3-киназы класса 1, построенные из каталитической субъединицы 110 кДа и адапторной субъединицы р85. В инсулинзависимых тканях найдены две формы каталитической субъединицы - р110a и р110b. Известны также две формы адапторной субъединицы, р85a и р85b, с высокой гомологией аминокислотной последовательности и пространственной структуры: каждая из них содержит два SH-домена, два пролин-богатых домена, SH3-домен и Bcr-гомологичный домен. Кроме того, обнаружено несколько укороченных форм адапторной субъединицы: они не содержат SH3-домена, Bcr-гомологичного домена и одного из пролин-богатых доменов. В адипоцитах человека экпрессируется только адапторная субъединица р85a, в то время как в клетках скелетных мышц 7 вариантов: р85a, р85b и пять укороченных. Разные варианты адапторных белков не в одинаковой мере включаются в фосфотирозиновые комплексы при инсулиновой стимуляции. Эти результаты указывают на возможность разветвления сигнального пути вследствие избирательной мобилизации определенного варианта адапторного белка (или определенного набора адапторных белков) в зависимости от природы первичного стимула.
Белки Ras входят в суперсемейство малых ГТФ-связывающих белков. Это небольшие белки (молекулярная масса 21 кДа, около 190 аминокислотных остатков), содержащие на С-конце ковалентно связанный фарнезильный или геранильный остаток:
С помощью такого гидрофобного конца белки Ras (p21ras) прикрепляются к внутренней поверхности плазматической мембраны. Ras вовлечены в разнообразные клеточные процессы, включая везикулярный транспорт, функции шаперонов, пролиферацию.
Как и все ГТФ-связывающие белки, Ras регулируются циклом ГТФ-белок (активная форма) U ГДФ-белок (неактивная форма). В этих превращениях участвуют еще и другие белки: GAP (GTPase activating factor), GEF (GTF exchange factor) и SOS; два последних белка обеспечивают отделение ГДФ от Ras и присоединение ГТФ.
В покоящейся клетке р21ras находится преимущественно в неактивной ГДФ-форме. Стимуляция клетки инсулином (а также другими факторами роста и митогенами) приводит к быстрому возрастанию количества активной ГТФ-формы. Происходит это следующим образом. Небольшой цитозольный белок Grb-2 (growth factor receptor bound protein), содержащий SH2- и SH3-домены, может нековалентно присоединяться к фосфорилированному РИ в области определенных фосфотирозиновых остатков.
Этому взаимодействию способствует один из субстратов РИ, а именно Shc. Далее образовавшийся комплекс взаимодействует с другим комплексом, содержащим белок Ras (р21ras). Белки Grb2 и Shc называют также адапторными белками, поскольку они связывают тирозинкиназные рецепторы (в данном случае РИ) с белком Ras. В комплекс также включаются белки, обеспечивающие обмен ГДФ/ГТФ и активацию Ras (SOS, GAP, GEF, OST). На цитоплазматической части рецептора инсулина образуется большая гроздь взаимодействующих белков. Таким образом инсулин активирует белок Ras, и инсулиновый сигнальный путь соединяется с сигнальным путем Ras.
Активация Ras является конечным звеном трансмембранной передачи сигнала и начальным звеном цитоплазматических и ядерных сигнальных путей. Эти пути составляют каскад протеинкиназных реакций с участием протеинкиназы Raf-1, МАПКК (митогенактивируемой протеинкиназы киназа) и МАПК (митогенактивируемые протеинкиназы). Активированный Ras приобретает способность соединяться с протеинкиназой Raf-1. Raf-1 находится в цитозоле в соединении с некоторыми белками теплового шока, и в этом состоянии не обладает протеинкиназной активностью; фермент активируется в результате соединения с белком Ras. Этот процесс сложен: для полной активации Raf-1 требуется его присоединение к плазматической мембране, фосфорилирование по тирозиновым остакам ферментом саркокиназой (Src), фосфорилирование по сериновым и треониновым остаткам специфической протеинкиназой С, а также взаимодействие с рецептором инсулина. Таким образом, в этот момент гроздь белков на рецепторе инсулина еще больше вырастает.
Активированная протеинкиназа Raf-1 фосфорилирует (активирует) МАПКК, которая фосфорилирует МАПК. Активированная МАПК фосфорилирует определенные белки цитоплазмы (в частности - протеинкиназу pp90S6, фосфолипазу А2 и рибосомальную киназу).
Сигнал может передаваться также и в ядро, обеспечивая регуляцию транскрипции определенных генов: фосфорилированная МАПК фофорилирует (активирует) ряд факторов транскрипции.
Путь Ras активируется не только инсулином и его рецептором, но и многими другими гормонами, факторами роста и их рецепторами. С этими процессами, в частности, связана клеточная пролиферация и трансформация. Однако конечный ответ клетки на разные сигналы бывает различным, оказывается специфичным для данного первого вестника сигнала. Это связано, в частности, с наличием вариантов белков (семейства), участвующих в трансдукции сигнала.
Активация пути Ras инсулином приводит, наряду с другими ответами клетки, к изменению обмена гликогена:
Одним из ферментов каскада протеинкиназ, активируемых комплексом Ras, является протеинкиназа рр90S6. Этот фермент катализирует Сер/Тре-фосфорилирование протеинфосфатазы, связанной с гранулами гликогена (ПФГр-1). Фосфорилированная (активная) форма ПФГр-1-Р дефосфорилирует (активирует) гликогенсинтазу (ускоряется синтез гликогена). ПФГр-1-Р дефосфорилирует также киназу фосфорилазы и гликогенфосфорилазу (прекращается мобилизация гликогена). Таким длинным путем инсулиновый сигнал доходит до одного из конечных, эффекторных звеньев.
Действие инсулина начинается с процесса его соединения с a-субъединицей рецептора. Образование инсулинорецепторного комплекса – основной момент в дальнейшем проявлении многочисленных биологических эффектов инсулина. Связывание инсулина с рецептором приводит к его самофосфорилированию с участием рецепторной протеинкиназы, что происходит до или в период поглощения инсулинрецепторного комплекса. Активированный таким образом рецептор с участием фосфолипазы С способствует гидролизу мембранных фосфолипидов (гликозилфосфатидилинозитол), сопровождающемуся образованием инозитолтрифосфата и диацилглицерина. Активированный рецептор запускает цепочку последовательного фосфорилирования других белков, включая серинкиназную активность. Он, возможно, также взаимодействует с ГТФ-связывающими белками или цАМФ, приводя к активированию фосфорилирования/дефосфорилирования, стимулирует фосфодиэстеразу, снижает активность протеинкиназы, результатом чего и является изменение функции мембраны клетки.
Одновременно с этим процесс внедрения в клетку инсулинорецепторного комплекса влияет на эндоплазматический ретикулум, активируя рециркуляцию белков-транспортеров глюкозы внутрь клетки. Этот же комплекс взаимодействует с микросомами, лизосомами и ядерными структурами. После диссоциации рецептор возвращается на мембрану клетки, а инсулин активирует процессы дефосфорилирования ядерных белков, изменяет обмен мРНК, приводя к увеличению синтеза белков и другим “поздним” эффектам биологического действия инсулина.
Поиски механизмов нарушения пострецепторного действия инсулина не увенчались успехом [Перцева М.Н., 1996], но показали высокую экономичность и пластичность взаимодействия инсулина с тканями. Действие инсулина на клетки не требует синтеза вторичного посредника, некоторые метаболические эффекты инсулина могут реализовываться и без участия тирозинкиназного каскада, часто одна и та же молекулярная стратегия используется для достижения разных эффектов.





